Extending the theory of sexual selection
Sexual selection is a powerful evolutionary force that underlies the evolution of exaggerated male characters, such as horns and other weapons used in male-male competition for access to females, or conspicuous songs and displays used to attract and persuade females into mating. Importantly, sexual selection results from a co-evolutionary interplay between the sexes: male ornaments evolve to match a female's preference, and a female preference in turn evolves as a function of the variation in male phenotypes. This evolutionary interdependence of male and female characters makes it difficult to make verbal predictions. In such a case, formal models are an ideal tool to generate logically consistent hypotheses. In a recent review (Kuijper, Pen & Weissing, Ann. Rev. Ecol. Evol. Syst., in press), we tallied more than 150 recent models on the co-evolution between the sexes.
Still, many aspects of sexual selection are not yet well understood. For example, many seemingly general predictions of sexual selection theory are based on the implicit assumption that male and female traits will converge to equilibrium. However, co-evolutionary processes like sexual selection – which in many respects resembles an arms race between the sexes – do often not converge. As shown by Van Doorn & Weissing (Am. Nat. 2006), such non-equilibrium dynamics can have major implications, e.g., on the diversity and the information content of sexual signals.
Moreover, there are many areas where sexual selection theory needs to be extended. For example, mutual mate choice or cryptic female choice have not yet received much attention by modellers. One field with major gaps in our knowledge is sexual selection in hermaphrodites. Indeed, virtually all sexual selection models were developed for species with two separate sexes. This is surprising, since hermaphroditism is the dominant mode of reproduction in plants, while in animals (where about 6% of all species are hermaphroditic) some of the most spectacular examples of sexual conflict have been found in hermaphrodites. We therefore developed a variety of sexual selection models that were specifically tailored to (simultaneous) hermaphrodites.
It is easy to realize that not all results of standard sexual selection theory can directly be extrapolated to hermaphrodites. First, standard theory assumes that the expression of sexual ornaments is sex-limited, implying that only males have to bear the costs of such ornaments. Since such sex-linkage is impossible in (simultaneous) hermaphrodites, the costs of ornamentation are higher than in species with two separate sexes. Second, standard theory assumes standard sex roles, with one choosy sex and the other sex competing for being chosen. In hermaphrodites, the complications of mutual mate choice cannot be avoided. Third, selection on males and females often acts in opposing directions. For example, males are typically selected to realize as many matings as possible, while a much smaller number of matings is optimal for females. In hermaphrodites, this sexual conflict translates into a genetic conflict between the male- and female-function genes of one and the same individual. Fourth, sexual selection in hermaphrodites is intricately related to sex allocation, i.e., to the relative investment into male and female function. In a first model (Van Velzen, Schärer & Pen, Proc. R. Soc. B, 2009), we studied the interaction of sex allocation and sperm competition. We showed, for example, that the mechanism of cryptic 'female' choice can have important implications for the outcome of evolution and that sexual conflict in hermaphrodites can trigger the evolution of separate sexes (more precisely, into males and into hermaphrodites with female-biased sex allocation).
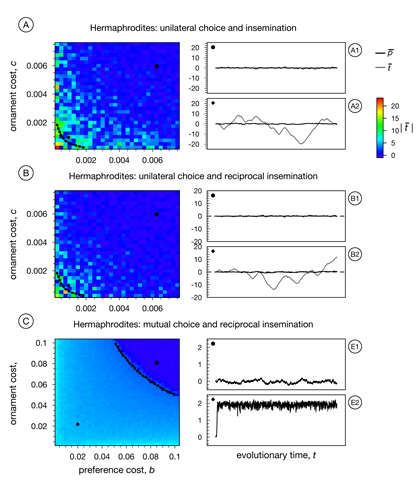
By constructing models in a systematic and step-by-step fashion and by comparing the outcome of individual-based simulations with analytical predictions, we are building up an inventory of sexual selection models allowing us to evaluate relative importance of ingredients like reproductive mode (hermaphrodites versus gonochorists), mating system (e.g., unilateral versus mutual mate choice), or timing of mate choice (pre- versus post-copulatory). This is illustrated by Figure 1, which shows the evolution of costly preferences in hermaphrodites. While sexual selection gets most easily off the ground in case of mutual mate choice, the strength of sexual selection (indicated by the degree of evolved ornamentation) is highest in case of unilateral mate choice. The outcome is hardly affected by the directionality of insemination. Comparing this figure with a comparable figure for gonochorists leads to the conclusion that – at least in the case of mutual mate choice – sexual selection may well be as strong in hermaphrodites as in organisms with separate sexes; something regarded as impossible by many evolutionary biologists, including Charles Darwin. Moreover, our study also demonstrates that simple models of sexual selection are not very robust when it comes to particularities of how individuals choose their mates.
| Last modified: | 03 December 2015 12.54 p.m. |