The evolution of animal 'personalities'
In many animal species, ranging from squids to chimpanzees, individuals of the same sex, age and size differ consistently in whole suites of correlated behavioural tendencies, comparable to human personalities. Various ‘behavioural types’ coexist within the same population, and such types exhibit behavioural consistency over time (e.g., types that are more aggressive as juveniles are also more aggressive as adults) and across contexts (e.g., types that are more aggressive towards conspecifics are also bolder towards predators). Both types of consistency indicate limited behavioural plasticity to a degree that, from an adaptive point of view, is often surprising. Shouldn’t a more flexible structure of behaviour provide a selective advantage? On a proximate level, behavioural consistency can often be understood in terms of the architecture of behaviour, that is, the genetic, physiological, neurobiological and cognitive systems underlying behaviour. But the question remains why architectural constraints on the flexibility of behaviour have not been removed by natural selection, and why not a single behavioural type outcompetes all other types in the population.
We developed a series of conceptual evolutionary models to address these questions. With these models we aim to address aspects of animal 'personalities' that appear to have some universality (i.e., occur in a range of animal species). In a first attempt (Wolf et al., Nature, 2007), we showed that individuals differing in future fitness expectations (e.g., individuals having territories of different quality) should systematically differ in their risk-taking behaviour. In short, individuals with high fitness expectations (having 'much to lose') should be more risk-averse in all kinds of contexts than individuals with low fitness expectations. This 'asset protection principle' does not only provide an explanation for the commonly observed boldness-aggressiveness syndrome, but also allows to make a priori predictions on the 'architecture' of risk-related behaviour. Recently, our predictions were confirmed in experiments with various organisms (including insects, fish, birds and mammals).
In a second model (Wolf et al., PNAS, 2008), we provided an explanation for the common observation that individuals often differ along a responsiveness (reactivity, flexibility, awareness) axis: at the unresponsive end of this spectrum we find individuals that quickly develop routines and show relatively rigid behaviour; at the responsive end we find individuals with a more extrinsic control of behaviour; these individuals are more flexible and readily respond to environmental stimuli. We could demonstrate that selection on responsiveness is negatively frequency dependent. This explains the evolutionary emergence of behavioural types differing in responsiveness. However, our model also partially explained why individuals should be consistent in their responsiveness over time and across contexts.
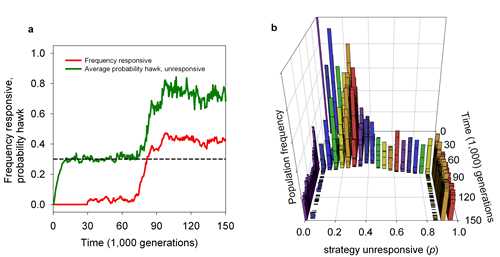
This aspect is address in a third model (Wolf et al., Proc. R. Soc. B, 2011), which studies responsiveness in the context of social interactions (also called 'social sensitivity'). We could show (Figure 1) that consistent differences in responsiveness can evolve through a co-evolutionary feedback process. The model reveals that responsive strategies can get off the ground whenever there is sufficient variation in the population (e.g., due to mutation and drift), since responsive individuals can exploit small deviations from equilibrium. Subsequently, the presence of responsive individuals triggers the evolution of behavioural consistency in unresponsive individuals, since – in two-strategy games with a mixed-strategy ESS – consistent individuals are exploited to a lesser extent than inconsistent ones. Behavioural consistency makes individuals predictable and, hence, favours responsiveness. In other words, there is a positive feedback between behavioural consistency (and predictability) and social sensitivity. We currently investigate the implications of this feedback for the evolution of cooperation.
In an invited review (Wolf & Weissing, Phil. Trans. R. Soc. B, 2010), we made an inventory of the most important evolutionary mechanisms leading to individual variation in behaviour and to consistency of behaviour, both in time and across contexts. In a second invited review (Wolf & Weissing, Trends Ecol. Evol., in press), we discuss the ecological and evolutionary implications of individual variation in behaviour.