Multispecies resource competition
In previous years, models produced within the Theoretical Biology group (in close collaboration with Prof. Jef Huisman, University of Amsterdam) have shown that the dynamics of interspecies competition is much more complicated than ecological textbooks seem to suggest. As soon as species compete for more than two limiting resources, the dynamics of competition can easily lead to oscillations and chaos. This non-equilibrium behaviour has important implications. Most importantly, it allows the stable coexistence of many competitors on a small number of limiting resources, whereas at equilibrium the number of coexisting species never exceeds the number of limiting resources. In fact, we could show that – away from equilibrium – hundreds of competitors can stably coexist on only three limiting resources.
The main focus of this project is the study of multispecies competition when nutrients can be stored by the competing organisms. Models including nutrient storage are more realistic but theoretically much more intricate than the standard models of resource competition. It is not self-evident whether storage enhances or limits the tendency for non-equilibrium dynamics. By means of a combination of analytical and numerical techniques, we could show that competition models with nutrient storage display the same dynamical behaviour (including oscillations, chaos and the coexistence of many species on few resources) as the simpler models without storage. However, there were also some notable differences in the dynamics of competition. In particular, specific trade-offs in the species consumption policies induced equilibrium behaviour while non-equilibrium dynamics would have resulted in systems without storage.
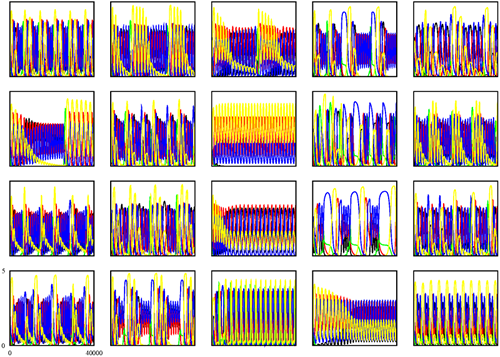
In order to achieve a better understanding of the factors shaping the dynamics of competing species, we performed a systematic comparison between various classes of competition models (Lotka-Volterra, Monod, Droop). We could demonstrate that some general rules (such as the well-known requirement for species coexistence, "intra-specific competition must be more intense than inter-specific competition") can be extrapolated from simple to complex models. In particular, the main mechanism behind competitive oscillations is the same in all models: the existence of a non-transitivity in the competitive hierarchy. For the majority of parameter combinations, complex models like the Droop model behave in a very similar way as the much simpler Monod model (Fig 6). However, as indicated in Figure 7, neglecting the complications generated by including nutrient storage can also lead to highly misleading conclusions.
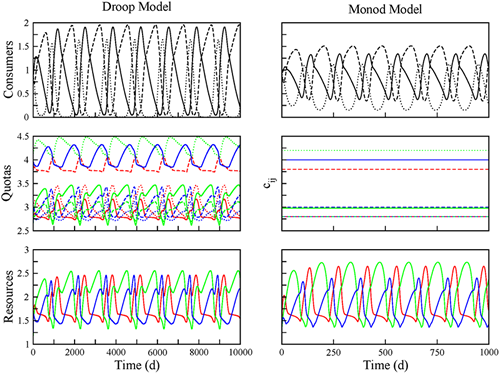
On the one hand, the robustness of model outcomes with respect to model assumptions is in line with line for a niche based perspective of community ecology, in which diversity is the consequence of the species properties. However, the addition of more mechanistic detail, like the introduction of nutrient storage, reveals that predictability becomes rather difficult, if not pointless, because small variations in biological parameters or initial conditions can lead to equilibrium coexistence, nonequilibrium coexistence, chaotic dynamics, or competitive exclusion (Fig 7). The implication of these contrasting results is that competitive communities may behave in a consistent way from a statistical perspective, but not on a case-by-case basis.
In a separate project, we investigated how non-trophic interactions can modify the fate of competition. Together with Ciska Veen (COCON) we studied the interplay between competition, mutualism and parasitism using a well-known model of plant-soil feedback. In this model, competition between two plants A and B is affected by the state of the soil community. Using graphical methods we determined that if the soil feedbacks are mostly negative (e.g. parasitism), competitively dominant plants become invadable, enhancing coexistence. An interesting consequence is the emergence of competitive oscillations between two species, in contrast with the minimum of three species required in standard models (Lotka-Volterra, Monod, Droop). If the soil feedbacks are mostly positive (e.g. mutualism), coexistence via mutual invasion is less frequent, although alternative coexistence states may be a possibility.