Life-history variation and the evolution of animal personalities
In recent years evidence has been accumulating that personalities are not only found in humans but also in a wide range of other animal species. Individuals differ consistently in their behavioural tendencies and the behaviour in one context is often correlated with the behaviour in multiple other contexts. Some great tits, for example, are consistently more aggressive than others, and aggressive birds differ from non-aggressive birds in many other respects like foraging behaviour or the exploration of novel environments. At present, the existence of such personalities is puzzling in several respects. First, why do different personality types stably coexist? Second, why is behaviour not more flexible but correlated across contexts and through time? And third, why are the same type of traits correlated in very different taxa? We aim at developing models to understand these kinds of questions better. We believe that there is not a unique explanation for animal personalities. This project therefore follows a pluralistic approach by investigating several different explanatory frameworks for the evolution of animal personalities. Here we summarize results of one of these subprojects.
We developed a theory that provides a link between life-history variation and animal personalities. This work was inspired by the observation that some of the most prominent personality traits described in the literature can be categorized in terms of risk-taking behaviour. A good example is the correlation between aggressiveness towards conspecifics and boldness towards predators: individuals that take more risk in intraspecific fights also take more risk when confronted with a predator. This aggression-boldness syndrome has been described for many species including fish, birds and rodents. From life-history theory it is known that individuals should adjust their risk-taking behaviour to their residual reproductive value, i.e., their expected future fitness. Individuals with relatively high expectations should be relatively risk-averse, since they have to survive in order to realize those expectations. By the same reasoning, individuals with relatively low expectations should be relatively risk-prone since they have little to lose. Consequently, whenever individuals differ in their fitness expectations, we should expect stable individual differences and correlated behavioural traits: some individuals are consistently risk-prone whereas others are consistently risk-averse.
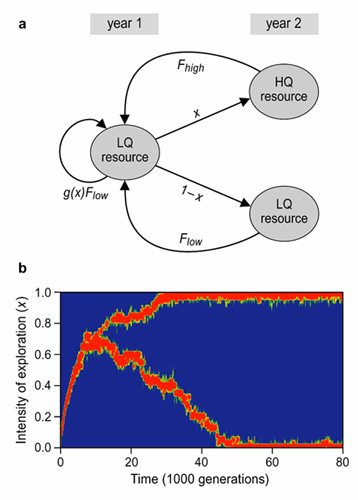
To investigate these intuitive arguments we studied a simple model (fig. 6a) in which individuals face a trade-off between current (i.e., year 1) and future reproduction (i.e., year 2) using a combined analytical and individual-based simulation approach. For illustrative purposes we assumed that this trade-off is mediated by exploration behaviour: the more thorough an individual explores its environment, the more emphasis it puts on future reproductive success. In a first step we showed that the trade-off between current and future reproduction can easily give rise to the stable coexistence of different life-history strategies: at the evolutionary equilibrium, some individuals explore their environment thoroughly, thereby investing in future reproductive success, whereas others explore superficially, putting more emphasis on current reproduction (fig. 6b).
We then extended this basic life history by assuming that each individual is confronted with a number of risky contexts throughout its lifetime. In particular, each individual may face one or more foraging decisions under predation risk (anti-predator games) and one or more aggressive encounters with randomly chosen conspecifics. In each of the anti-predator games an individual can either behave in a bold or a shy manner. Only bold individuals receive a payoff (in terms of higher fecundity) but they also incur some risk of dying. Aggressive encounters are modelled as hawk-dove games, where hawks receive a higher payoff than doves but do not always survive hawk-hawk interactions.
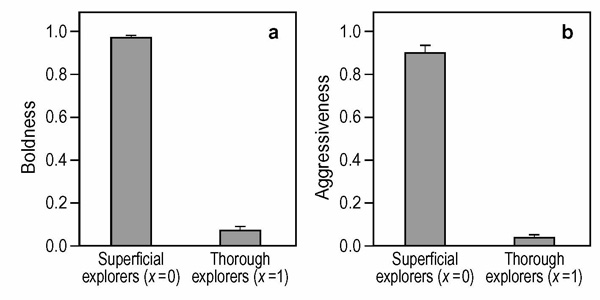
We first considered the most basic scenarios where individuals either play one anti-predator game or one hawk-dove game (fig. 7). Our analysis confirms the general principle that individuals with higher future expectations (in our case: thorough explorers) should behave in a more cautious way. At the evolutionary equilibrium, superficial explorers behave boldly (aggressively) whereas thorough explorers show low levels of boldness (aggressiveness).
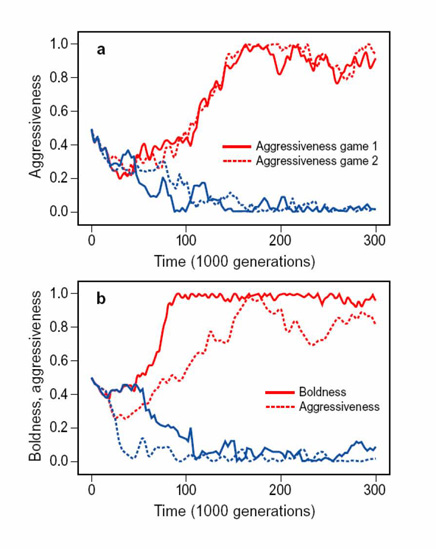
To show that the same principle gives rise to personalities we considered scenarios where individuals are confronted with several risky games throughout their life. We find that natural selection gives rise to stable individual differences within the same game and correlated behavioural traits across different games (fig. 8). Figure 8a considers the scenario where individuals play two hawk-dove games sequentially. Here superficial explorers evolve high levels of aggression in both games whereas thorough explorers evolve to be consistently non-aggressive. Figure 8b depicts the scenario where individuals face one anti-predator and one hawk-dove game sequentially. Here evolution gives rise to the coexistence of superficial explorers that are both bold and aggressive and thorough explorers that are shy and non-aggressive. In other words, we robustly observe the well-known behavioural syndrome linking aggression and boldness, which has been reported for numerous species in a diversity of taxa.
Up to this point we assumed, for simplicity, that individuals reproduce asexually. We obtain qualitatively the same results for scenarios that allow for recombination and diploid genetics, as long as individual alleles have a large phenotypic effect. Under this condition disruptive selection gives rise to a small number of discrete phenotypes, each corresponding to a distinct personality type. A more realistic approach, however, would be to consider quantitative traits, which are often thought to be influenced by many loci with small effects. We therefore incorporated such quantitative genetics into our model.
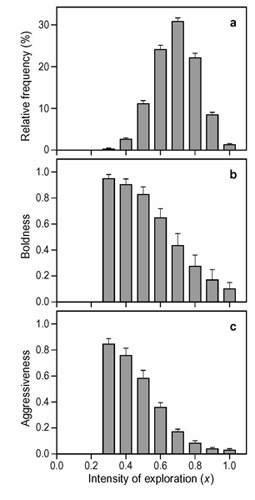
Figure 9 considers a scenario where individuals face one anti-predator and one hawk-dove game sequentially. As shown in figure 9a, disruptive selection does now not result in two extreme exploration (i.e., life-history) strategies but in the stable coexistence of a broad range of explorative behaviours. Similarly, with respect to both boldness and aggressiveness, we find a gradation of behavioural traits ranging from low to high levels (fig. 9b, c). A clear pattern emerges: the more superficially an individual explores its environment (fig. 6a), the more boldly it behaves in the anti-predator game (fig. 9b) and the more aggressive it is in the hawk-dove game (fig. 9c). In other words, in the case of quantitative genetic variation we find a whole spectrum of personality types, which is in line with many empirical studies.
In summary, our model offers a plausible explanation for individual differences within a population, the evolution of behavioural correlations within and across contexts and the fact that particular traits like boldness and aggressiveness tend to be associated. Our theory is well testable by predicting (1) under what circumstances behavioural correlations should occur, (2) what particular traits should be correlated and (3) what sign the correlations should take. Behavioural correlations are to be expected whenever individual differences in residual reproductive value (i.e., expected future fitness) occur. One might think of the difference between high- and low-ranking individuals in a dominance hierarchy, between dispersers and philopatric individuals in a metapopulation or between residents and floaters in a territorial system. In any such situation we would expect correlations between those behavioural traits that involve risks which might prevent individuals from reaping the returns from reproductive investments. In addition to intraspecific aggression and boldness one might also think of behaviours such as brood defence, nest guarding or conspicuous displays to attract mates. All other things being equal, we would expect such traits to be positively correlated.